Pseudorabies in Swine
Definition
Pseudorabies (PRV) is a major viral disease manifested in swine by signs and lesions that vary among different age groups. The disease is characterized by three overlapping syndromes that reflect lesions in the central nervous system (CNS), respiratory system or reproductive system.
Occurrence
Among domestic animals, swine are the only natural host of pseudorabies virus (PRV). All age groups not previously exposed or vaccinated are susceptible. The disease was eradicated from the US commercial pig industry in 2004 but remains in some localized feral swine populations. The disease remains common in many other major swine-raising countries. Most domestic animals (cattle, sheep, dogs, cats, and goats but not horses) and many wild animals (rats, mice, raccoons, opossums, rabbits, and several fur-bearing mammals) are susceptible to the virus but transmission only occurs when these species are kept in close contact with acutely infected swine; death is the usual outcome in these aberrant hosts. Pseudorabies occurs with some frequency in both cattle and sheep, especially those in close contact with swine. There is no evidence that PRV is a health threat to humans.
Historical information
The disease is named after the Hungarian veterinarian Dr. Aladár Aujeszky who linked the disease in cattle, dogs, and cats in 1902. Pseudorabies was not identified as a viral disease in swine until 1909.
Prior to 1960, the disease in swine was important in Eastern Europe but major outbreaks did not occur in the US until the mid-1970s. Incidence of the disease increased with the widespread introduction of total confinement and continuous farrowing during this period. Additionally, antiserum used in control of classical swine fever prior to this period often contained antibodies to PRV and likely contributed to suppression of clinical signs on many farms. Active PRV eradication programs exist in many countries and have been successful, in large part due to efficacious differential vaccination.
In 1989, the US embarked on a 5-stage Federal/State/Industry program for eradication of PRV in swine; eradication of PRV from the commercial industry was achieved in 2004.
Etiology
Pseudorabies virus (PRV) is an alphavirus in a subfamily of Herpesviridae. Isolates vary in virulence and pathogenicity. Like many other herpesviruses, PRV often persists in a latent state in recovered animals. The virus is only moderately resistant outside the host, survival depending largely on environmental factors. Except during cold weather, virus probably does not survive more than two weeks outside the pig. Under exceptional conditions it may survive longer in infected fetuses, dried tissue, and buildings. The virus can be destroyed by many disinfectants, including orthophenylphenol, quarternary ammonium or iodine compounds, and 5% sodium hydroxide.
At least nine proteins, many of them structural proteins on the virus envelope, have been identified. By genetic engineering it has been possible to prepare effective vaccines that omit one or more of the proteins. Antibodies to such vaccines differ sufficiently from antibodies to wild virus so that serologic tests can identify naturally infected herds. Protection induced by commercial PRV vaccines appears to be effective against all wild types of the virus.
Epidemiology
Pseudorabies virus is spread and persists by several mechanisms. Swine that recover from PRV excrete large amounts of virus in saliva and nasal secretions, and perhaps in urine and feces, for up to two weeks. Virus can persist in the tonsils of carrier swine for at least several weeks. Latent virus can persist in the CNS for many months. Recrudescence and shedding of virus often occur after stress.
Inapparent shedders frequently are the means of introduction of virus into susceptible herds. Once introduced, the virus spreads by nose-to-nose contact, through feed and water contaminated by oral secretions and by aerosols coughed into the air. Circumstantial evidence suggests that an infectious aerosol can spread several miles.
In a farrowing house, aborted fetuses or infected piglets born alive or incubating the disease soon contaminate the house. Virus may survive long enough to infect piglets of the next farrowing.
Feral swine remain a possible source of infection to domestic swine. Other PRV-infected animals (e.g. rats, mice, dogs, cats, raccoons, opossums) are considered “dead-end” hosts and will shed PRV only a short time. They may, however, visit swine facilities and either shed virus that contaminates the facility or they may die and be eaten by swine. The possible role of flies or birds in transmission is dubious.
Pathogenesis
Pathogenesis is variable, depending on virulence of the viral strain, age of the pig and the route and severity of exposure. Experimentally, pigs can be infected by many routes of inoculation. Under field conditions the oral-nasal routes of exposure probably are most common. Virus often localizes in epithelium of the tonsils. The primary site of replication is epithelium of tonsils and nasopharynx. With highly virulent strains of virus there may be a short period of viremia with dissemination to many tissues. Most viral strains have a tropism for the CNS and upper respiratory tract.
The virus also can spread from a primary site of infection along nerves to the CNS, then to the spinal cord and spread centrifugally along nerve trunks. Virus often localizes in ganglia, e.g. the trigeminal nerve ganglia and paravertebral ganglia, and may become latent there. It may be reactivated later by stress.
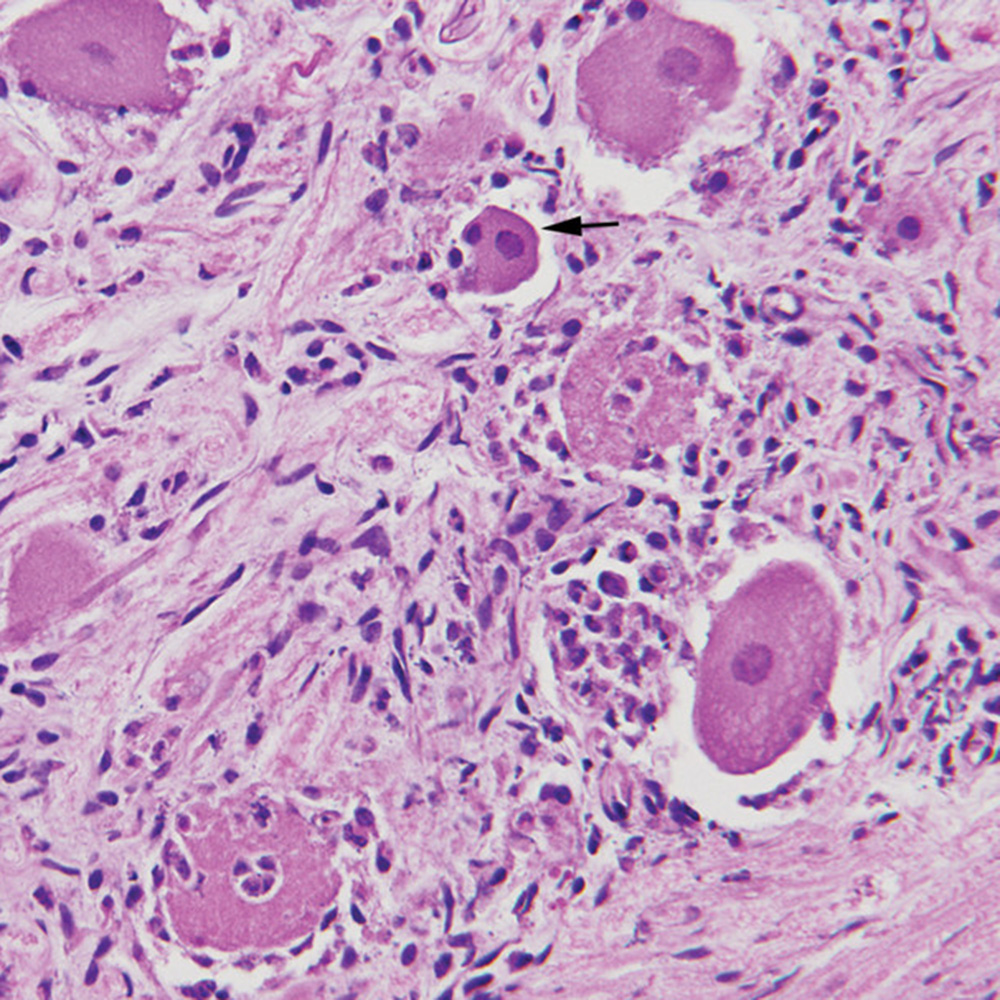
In many areas of localization, including respiratory epithelium, the virus causes inflammation and necrosis of tissue. In some organs and tissues, necrotic areas often are large enough to be grossly visible. Virus may cross the placenta to infect some or all fetuses. In the reproductive tract, the virus causes endometritis, vaginitis, and necrotic placentitis. Lesions and virus may be present in infected fetuses. Intranuclear inclusion bodies are produced in a variety of cells in swine of all ages, including fetuses.
Clinical signs
Signs vary, depending largely on the immune status of the dam and the age of the pigs affected. In general, younger pigs are more severely affected but become more resistant as they age. Neonatal pigs born to naïve sows are highly susceptible to infection and piglets become ill two to four days after exposure. Very young neonates may die without signs even having been noticed. Nursing pigs usually show signs of neurologic involvement. Signs may include high fever, depression, anorexia, tremors, incoordination, dog-sitting position, vomiting, foaming at the mouth, blindness, paddling, coma and convulsions. Death often occurs within one to three days. Morbidity and mortality may approach 100% in neonates, especially in naïve herds. Piglets nursing recovered or vaccinated dams are well protected from clinical signs and infection with the virus.
Weaned pigs (three to nine weeks of age) have similar signs but mortality is usually much lower. Respiratory signs often occur in the older pigs of this age group. They include sneezing, nasal discharge, sometimes coughing and labored breathing. Most of the pigs survive, provided they do not succumb to secondary infections, especially bacterial pneumonia.
In susceptible, grow/finish swine (ten weeks of age to market weight), respiratory signs predominate and morbidity is high. Signs include a febrile response, depression, anorexia, sneezing, coughing and nasal discharge. CNS signs occur in occasional pigs and vary in severity from tremors to convulsions. Most pigs recover in about seven to ten days.
In breeding herds, clinical signs may not be apparent except during the period of initial infection. These signs are primarily respiratory and most animals will recover. In pregnant sows or gilts, reproductive failure sometimes occurs. Dams infected in the first trimester may absorb their fetuses and return to estrus. Those in the second or third trimester may abort or have mummified, stillborn or weak pigs. Those infected close to term may deliver infected piglets that die within a few days. Reproductive failure usually has a modest incidence of < 20%.
Lesions
Neonates that die acutely after infection may have no gross lesions. In piglets showing CNS signs, congestion of the meninges often is visible. In those showing respiratory signs, there may be a rhinitis with patchy epithelial necrosis. The rhinitis can be seen after splitting the skull and nasal bones midsagittally. The inflammation often extends into the fauces and larynx, and occasionally into the trachea or esophagus. Other gross lesions may include keratoconjunctivitis and necrosis of the tonsils. Gross and/or microscopic foci of necrosis may be seen in the liver, spleen, lungs, adrenals and lymph nodes. Similar foci sometimes are present in aborted fetuses.
Microscopic lesions often include a panencephalitis, ganglioneuritis at multiple sites, and perhaps nonsuppurative meningitis. The most severe lesions in the brain usually are in the cerebral cortex and often include neuronal degeneration and necrosis. In the lungs there may be bronchitis, bronchiolitis and alveolitis with hemorrhagic foci of necrosis but lung lesions generally are neither severe nor specific. Intranuclear inclusion bodies may be found at many sites, often within neurons, astrocytes, oligodendroglia and respiratory epithelium. They are more easily demonstrated in mercury-fixed tissues.
In dams that have infected fetuses or abort, the uterine wall may be thickened by inflammatory edema. Necrotic placentitis may be apparent in the placenta of infected fetuses and neonatal piglets.
Diagnosis
Diagnosis often can be made on the basis of prevalence of the disease in the area, history, signs, gross and microscopic lesions or serologic testing. Serologic tests available include serum neutralization (SN), latex agglutination (LA), and enzyme-linked immunosorbent assay (ELISA). SN is the standard regulatory test but the LA and ELISA tests are widely used as screening tests. Serologic testing is widely used for herd diagnosis. Accurate serologic tests that can differentiate vaccination antibody from wild type are widely available and are commonly used as part of national disease control programs. A fluorescent antibody test on tissue sections, immunohistochemistry on formalin-fixed tissues, or virus isolation may be used to identify virus in the brain, tonsils, and spleen. A polymerase chain reaction (PCR) test has been described but is not in common use.
Control
Pseudorabies is currently considered an exotic viral disease in the US since its eradication in 2004. In countries where the disease remains endemic, it may be possible to prevent exposure of a negative herd to PRV through strict quarantine and isolation procedures. Only seronegative animals should be added, as necessary, to a herd. Additions should be made only after a quarantine period of at least one month and negative serologic tests before and after quarantine. Cats, dogs, rats, mice and all wildlife should be prevented from coming in contact with swine.
An all in/all out system with cleaning and disinfection between farrowings is helpful. Gene-deleted modified-live vaccines are available and their use is accompanied by the application of ELISA serologic tests that can differentiate infected from vaccinated swine. These vaccines are safe and concerns about return to virulence have been effectively dismissed. Pseudorabies vaccines increase resistance to PRV infection, virtually eliminate clinical signs associated with infection, and minimize the amount of viral shedding that results if infection occurs. The incidence of latency and frequency of recrudescence of virus in latently infected swine are also decreased in vaccinated swine.
There is no effective treatment for PRV-infected swine but weaned and older swine often recover.